
keywords: time crystal myelin
There is a moment in the history of philosophy where three great accounts of time converge without knowing it. Husserl describes the living present as a horizontal flow, retention holding the just-passed, protention reaching toward the about-to-arrive, consciousness stretched like a moving horizon across the surface of duration. Bergson describes duration as something deeper than flow, the whole accumulated past pressing up into the present moment, not as memory but as the thickness of now, time experienced not as a sequence of instants but as a continuous swelling of what has been into what is. And Heidegger, approaching from a different direction entirely, insists that Being and Time are not two things in relation but one thing seen from two angles: Dasein does not exist in time the way a stone sits in a field. Dasein is temporal through and through, constituted by its having-been, always already ahead of itself, thrownness and projection folded into the single event of existence.
Three philosophers, three accounts, three languages. And underneath all of them, waiting in the biology, a structure that is all three at once.
The myelin sheath is a time crystal.
What the sheath actually does
Myelin is not insulation.
This point has been made elsewhere in this series and it bears repeating here because the insulation metaphor closes the mind precisely where it needs to stay open. The myelin sheath is the biological substance of the self. It is the accumulated condition of lived experience, wrapped around the axon in a spiral structure that is simultaneously spatial and temporal, simultaneously the record of what has been and the condition of what is happening now.
The sheath is formed by an oligodendrocyte or Schwann cell extending a process around the axon and rotating, wrapping layer upon layer of membrane in a tight spiral. As each layer is laid down, the inner tongue, the leading edge of the wrapping process, is always the newest layer, pressing inward toward the axon, the most metabolically active, the most immediately adjacent to the signal. The oldest membrane is the outermost layer, the most compact, the most deeply sedimented, furthest from the axon but still structurally present, still held in place by the paranodal proteins, the PNP proteins that act as molecular velcro, binding the folded membranes into immediate contact with each other across the layers of the spiral.
The past does not recede in the myelin sheath. The accumulated condition presses outward from the innermost layer, each new layer of experience added to the outside of the existing structure, the most recent closest to the signal, the most ancient furthest away but never absent. The past is not archived behind a retrieval mechanism. It is folded into the structure, held there, coexisting with the act of myelination that is happening right now.
Husserl’s horizon and Bergson’s depth
Husserl described the living present as a temporal structure with two dimensions. Retention is the horizontal backward reach: the note just played is still present in the hearing of the note playing now, not as memory but as the trailing edge of the present moment itself. Protention is the horizontal forward reach: the note about to be played is already shaping how the current note is heard, not as anticipation but as the leading edge of the present moment. Consciousness is not a point moving along a line of time. It is a moving horizon, thick with the just-having-been and the about-to-arrive, a living present stretched across time in both directions simultaneously.
This is the horizontal dimension of the myelin sheath. The act of myelination is not a point event. It is a process with retention and protention built into its structure. The axon firing now is the current note. The myelinated layers already laid down are the retention, the just-having-been that is still structurally present in the current event. The oligodendrocyte still wrapping, still extending its inner tongue, is the protention, the about-to-be-laid-down already shaping the structure of the coupling that is occurring now.
But Husserl’s horizontal account, however precise, does not capture the full depth of what the sheath does. For that we need Bergson.
Bergson’s duration is not the flow of time along a surface. It is the accumulation of time as thickness. The present moment is not thin. It carries the whole past within it, not as a series of archived moments retrievable on demand, but as the accumulated weight of everything that has been, pressing into the present and shaping what is possible now by virtue of what has been. Duration is not experienced as a line. It is experienced as a swelling, a deepening, a thickening of the present with everything the organism has lived.

This is the vertical dimension of the myelin sheath. The layers of the spiral are not equivalent. The innermost layer is the newest, the most recently laid down, the most immediately adjacent to the firing axon. The outermost layer is the oldest, the most compact, the most deeply inscribed. Between them, every layer that was ever laid down in the life of this sheath, in the life of this organism in this experience, is present simultaneously, folded into the depth of the structure, pressing outward from the innermost layer through the accumulated history of the self. The past does not recede in the myelin sheath. It accumulates. It presses. It thickens the present from within.
Husserl gives us the horizontal continuity of the living present. Bergson gives us the vertical depth of accumulated duration. The myelin sheath has both dimensions at once, in the same structure, not as two separate features but as the single geometry of the spiral.
The spiral that unifies them
Here is where the geometry becomes decisive.
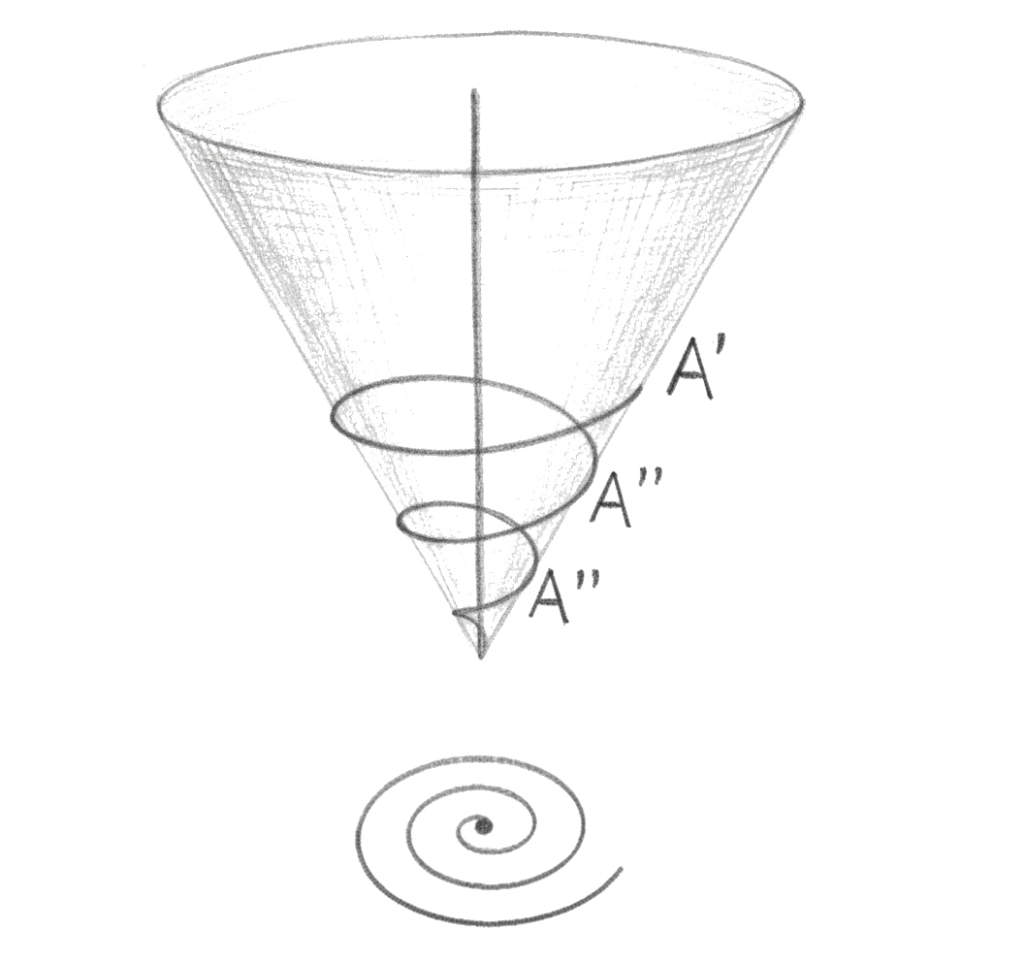
If time had only a horizontal dimension, it would be a line: past, present, future, sequential, irreversible, flat. If time had only a vertical dimension, it would be a column: depth upon depth, accumulation without forward movement, pure duration without direction. But the myelin sheath is neither a line nor a column. It is a spiral.
The spiral is the geometry that unifies horizontal and vertical without reducing either to the other. At every point on the spiral, the structure is simultaneously in horizontal relation to the adjacent turns, the retention and protention of the living present, and in relation to the layers inside and outside, the accumulated depth of duration pressing outward from the innermost layer. The spiral is not a rectangular lattice where horizontal and vertical are perpendicular and separate, meeting only at right angles. It is a helical totality where every point participates in both dimensions at once, where the horizontal flow and the depth of accumulated duration are not two properties of the structure but one property expressed simultaneously in two directions.
This is what Husserl and Bergson were each describing from one side. The horizontal phenomenologist and the philosopher of duration were both tracing the outline of the same biological structure without knowing it. The spiral myelin sheath is the biological realisation of what they could only approach from one direction at a time.
And the PNP proteins, the paranodal molecular velcro that binds the folded membranes into immediate contact, are what makes the structure resonant rather than merely structural. Without them, the layers would be adjacent but not coupled. The paranodal proteins bring past and present into immediate physical contact, not merely spatial proximity. They make the accumulated history of the sheath available to the present event of coupling, not as a retrievable archive but as an immediate structural presence. The whole past is available to the whole present because the paranodal proteins hold them together in the same vibrating structure.
Heidegger: Being and Time as one
Heidegger’s central claim in Being and Time is that the question of Being cannot be separated from the question of time. Dasein, the kind of being that asks about its own being, is not a thing that exists and also happens to be in time. Dasein is temporal through and through. Its having-been, its thrownness into a world it did not choose, is not behind it as past history. It is the condition from which every projection forward departs. Its being-toward-the-future is not ahead of it as an anticipated event. It is the structure of its existence right now, the way it is always already ahead of itself, reaching toward what it is not yet.
Being and Time are one because the being of Dasein is constituted by its temporality, and its temporality is constituted by the unity of having-been, present and coming-toward in a single structure that Heidegger calls care.
The myelin sheath is the biology of this unity.
The axon is Dasein. The firing of the axon is the present event of being-in-the-world, the encounter with what is here now. The myelinated sheath is the having-been, the accumulated condition of every previous encounter, folded into the structure and pressing outward from the innermost layer into the present event. The ongoing myelination, the inner tongue still wrapping, the newest layer still being laid down adjacent to the axon, is the being-toward, the projection forward, the structure of the sheath already reaching toward what it is not yet fully.
Being and Time, one thing. Axon and sheath, one thing. The event of firing and the accumulated condition of having-fired, one thing, not two.
Heidegger could not have known about PNP proteins. But the structure he was describing philosophically, the unity of thrownness and projection in the single event of care, is what the paranodal velcro achieves biologically. The past is not archived behind the present. It is held against it, in immediate structural contact, by molecular bonds that bring the whole having-been of the organism into the present event of coupling.
The cubic crystal and the resonance of selfhood
A crystal resonates because its molecular structure is periodic: the same arrangement of atoms repeating in all directions, creating a lattice that vibrates coherently when stimulated. A cubic crystal resonates isotropically, the same in all directions, because its periodicity is the same along every axis. The resonance is not a property added to the structure from outside. It is a property of the structure itself, a consequence of the way its constituents are arranged in relation to each other across the whole.
The myelin sheath resonates in an analogous sense, but the resonance it enables is not acoustic. It is conscious. The sheath resonates when the incoming grey matter signal, the axon firing, couples with the accumulated myelinated condition of the organism’s history. The quality of that resonance, what it sounds like from the inside, is what we call experience. The particular frequency at which this organism’s sheath resonates, in this moment, with this signal, is what we call selfhood.
No two sheaths resonate identically, because no two organisms have lived identically. The spiral structure of each sheath is the unique crystallographic record of a unique life. The Myelin Mind thesis is that consciousness arises at the coupling between signal and structure. But it is now possible to say something more precise: consciousness arises when an incoming signal resonates with a spiral time crystal that is the biological inscription of a particular being in a particular time.
Not a being in time. A being as time.
This is what Heidegger meant. This is what the myelin sheath is. Being and Time are one because the sheath, which is being, is also time: horizontal in its flow, vertical in its depth, spiral in its unity, crystalline in its resonance, and unique in the way that only a life, and not a database, can be unique.
What this means for the wired mind
The wired mind locates consciousness in the neuron, in the signal, in the firing. It treats the axon as the subject and the sheath as the support structure, the insulation, the irrelevant coating. On this account, time is the medium in which neural events occur. The brain processes information in time, and the experience of time is itself one of the things the brain processes.
The time crystal argument inverts this completely. Time is not the medium in which the sheath exists. The sheath is the biological form that time takes in a living organism. The neuron is not the subject of experience. The coupling of neuron and sheath is the subject of experience, and the quality of that experience is determined not by the signal alone but by the crystalline structure of the accumulated condition it encounters.
The wired mind cannot account for duration, for the thickness of the present, for the way the whole past presses into the current moment not as memory but as structural depth. It cannot account for the felt unity of having-been and being-toward in the single event of consciousness. It cannot account for why no two conscious experiences are identical even when the stimulus is the same, why the same cup looks different to the same person on different days, why the same piece of music heard at twenty and at sixty are not merely interpreted differently but experienced differently, felt differently, resonating differently against a sheath that has been changed by everything that happened in between.
The wired mind has a signal and a processor. The Myelin Mind has a signal and a time crystal. The difference is the difference between information and experience, between processing and being, between a computation that happens in time and a life that is time.
Deleuze, Derrida and the fold
Deleuze took his crystal-image from Bergson. The crystal-image in Cinema 2 is the moment where past and present become indiscernible, where the actual event and its virtual coexist in the same image, where you cannot tell which side is the thing and which is the mirror. The crystal for Deleuze is the point where time folds back on itself and shows its own structure.
The myelin sheath does not merely resemble this. It enacts it. The actual, the axon being myelinated right now, and the virtual, the already-myelinated layers folded and tucked underneath, coexist in the same biological structure in a state of genuine indiscernibility. The inner tongue wrapping inward is the actual. The outermost compact layers are the virtual, the accumulated condition that was never itself experienced as a present moment but that is nonetheless the condition for every present experience. This is not a cinematic image of time. It is time itself, biological and literal, folding back on itself in the paranodal contact between what is being laid down and what has already been laid down.
And here Derrida arrives uninvited. The already-myelinated is a past that was never present. No organism ever experienced the laying down of its own myelin as a present moment. The sheath accumulates below the threshold of consciousness, in sleep, in development, in the slow metabolic work of oligodendrocytes going about their business in the dark. The accumulated condition that constitutes the self is, in Derrida’s precise sense, a trace: the arche-trace, the condition of possibility for all experience that was never itself experienced, the origin that was never an origin, the past that was never a present.
The myelin sheath is the biological arche-trace. The self is constituted by a history it never lived as a present moment. What you are is the accumulation of what was never, as it happened, available to you as experience. The condition of your consciousness is a past that consciousness never touched.
The wired mind has no language for this. The Myelin Mind has nothing else.
Jack Parry is a philosopher, polyglot and biomedical animator at Swinburne University of Technology. He is the author of The Myelin Mind: The Genesis of Meaning.